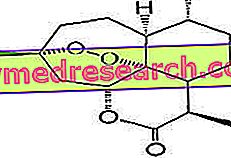
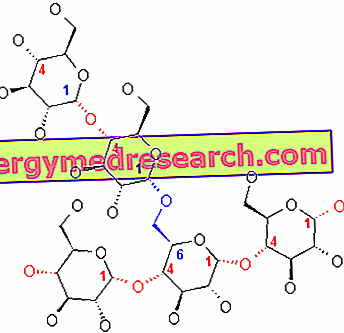
الجليكوجين هو جزيء ضخم (كتلة جزيئية من حوالي 400 مليون دالتون) من الجلوكوز ألفا حيث توجد بشكل رئيسي روابط α-1،4 glycosidic والتأثيرات في نسبة 1: 10 ، بسبب روابط α-1،6 glycosidic.
الجليكوجين هو مادة احتياطية ويتم تدهورها وإعادة تشكيلها باستمرار ؛ في كل كتلة الجسم الخلوية ، هناك حوالي 100 غرام من الجليكوجين: معظمها في الكبد حيث يكون متحركًا ، وبالتالي يمكن استخدامه كاحتياطي للأعضاء الأخرى (الجليكوجين في العضلات غير متحرك).
إن الإنزيمات التي تحفز تدهور وتوليف الجليكوجين كلها موجودة في السيتوبلازم ، لذا نحتاج إلى نظام ضبط يجعل غير نشط غير نشط عندما يكون الآخر نشطًا: إذا كان هناك توفر للجلوكوز ، يتم تحويل الأخير إلى جليكوجين (الأناقة) التي هي احتياطي ، والعكس بالعكس ، إذا كان هناك طلب على الجلوكوز ، ثم يتحلل الجليكوجين (الهدم).
إن الإنزيم الذي يشارك بشكل رئيسي في تدهور الجليكوجين هو فسفوريلاز الجليكوجين . هذا الإنزيم قادر على تشبيك رابطة α-1،4 glycosidic باستخدام ، كعامل ليتي ، orthophosphate غير عضوي: يحدث الانقسام بواسطة طريقة phosphorolytic ويتم الحصول على الجلوكوز 1-فوسفات.
في خمس أو ست وحدات من نقطة متفرعة ، لم يعد إنزيم الفوسفوريلاز الجليكوجين قادرًا على التصرف ، وبالتالي فهو ينفصل عن الجليكوجين ويتم استبداله بإنزيم إدماني وهو ترانسفيراز : في الموقع التحفيزي لهذا الإنزيم يوجد الهيستدين الذي يسمح بنقل ثلاث وحدات من السكريتيدات إلى أقرب سلسلة من الجليكوسيد (يهاجم الهيستدين الكربون الأول لجزيء الجلوكوز). الإنزيم الذي تم ذكره للتو هو glycosyltransferase . في نهاية عمل هذا الإنزيم ، تبقى وحدة غلوكوز واحدة فقط مع أول الكربون المرتبط بالكربون السادس لجلوكوز السلسلة الرئيسية. يتم تحرير وحدة الجلوكوز الأخيرة من السلسلة الجانبية من خلال عمل الإنزيم α-1،6 glycosidase (يشكل هذا الإنزيم الجزء الثاني من عترة الإنزيم) ؛ لأن النتائج في الجلايكوجين هي في نسبة 1:10 ، من الانهيار الكامل للجزيئ الماكرو نحصل على حوالي 90٪ من الجلوكوز 1-فوسفات وحوالي 10٪ من الجلوكوز.
يسمح عمل الأنزيمات سالفة الذكر بإزالة سلسلة جانبية من جزيء الجليكوجين. يمكن تكرار نشاط هذه الإنزيمات حتى يحدث تدهور سلسلة كاملة.
النظر في خلايا الكبد. يتم تحويل الجلوكوز (استيعابها من خلال النظام الغذائي) ، عندما يدخل الخلية إلى 6 الجلوكوز والفوسفات وهكذا يتم تنشيط. يتم تحويل الجلوكوز 6-فوسفات ، بسبب عمل فسفوغلوكوتاز ، إلى 1-فوسفات الجلوكوز: الأخير هو مقدمة غير فورية للتخليق الحيوي ؛ في عملية التخليق الحيوي يتم استخدام شكل من السكريات المنشط والذي يمثل بالسكر المرتبط بالثنائيات: عادة يوريدي ديفوسفات (UDP). ثم يتم تحويل الجلوكوز 1-الفوسفات إلى الجلوكوز UDP. هذا المستقلب يندرج تحت تأثير سينسيز الجليكوجين والذي يكون قادراً على ربط الجلوكوز UDP إلى نهاية غير قابلة للحد من الجليكوجين المتنامي: يتم الحصول على الجلايكوجين الممتد لوحدة غلوكوزيد و UDP. يتم تحويل UDP بواسطة nucleosidedifosfochinasi انزيم في UTP الذي يعود إلى الدورة الدموية.
يحدث تحلل الجليكوجين عن طريق عمل فسفوريلاز الجليكوجين الذي يحرر جزيء الجلوكوز ويحوله إلى 1-فوسفات الجلوكوز. في وقت لاحق ، يحول الفوسفوغلو كوموتاز الجلوكوز 1-الفوسفات إلى 6 الجلوكوز الفوسفات.
يتم تصنيع الجليكوجين ، قبل كل شيء ، في الكبد والعضلات: في الكائن الحي هناك 1-1.2 هكتوليتر من الجليكوجين موزعة خلال كتلة العضلات.
يمثل الجليكوجين في خلية عضلية احتياطي طاقة فقط لتلك الخلية في حين أن الجليكوجين الموجود في الكبد هو أيضًا احتياطي للأنسجة الأخرى ، أي يمكن إرساله ، مثل الجلوكوز ، إلى خلايا أخرى.
ثم يتم إرسال الجلوكوز 6 الفوسفات التي تم الحصول عليها في العضلات من تدهور الجليكوجين ، في حالة متطلبات الطاقة ، إلى تحلل. في الكبد ، يتم تحويل الجلوكوز 6-الفوسفات إلى الجلوكوز بفعل الجلوكوز 6-فوسفاتاز الفوسفات (خاصية أنزيمية لخلايا الكبد) ويتم توجيهه إلى تيار الدورة الدموية.
يعمل كلا من الجليكوجين سينسيز وفوسفوريلاز الجلايكوجين على وحدات الجليكوجين غير المختزلة ، لذا يجب أن يكون هناك إشارة هرمونية تتطلب تنشيط مسار واحد والكتلة الأخرى (أو العكس).
في المختبر كان من الممكن إطالة سلسلة الجلايكوجين ، واستغلال فوسفوريلاز الجليكوجين وباستخدام 1-الفوسفات الجلوكوز في تركيز عال جدا.
في الخلايا ، يحفز فوسفوريلاز الجليكوجين فقط رد الفعل التحلل لأن تركيزات الأيضات مثل لتحويل توازن التفاعل التالي إلى اليمين (أي نحو تدهور الجليكوجين):

نحن نرى آلية عمل فوسفوريلاز الجليكوجين: الأكسجين الأسيتال (الذي يعمل كجسر بين وحدات الجلوكوز) يرتبط بالهيدروجين الفوسفوري: يتم تشكيل تفاعل وسيط يعطى بواسطة carbocatione (على الجلوكوز الذي هو الحد الأقصى) الذي يرتبط به الفسفوريل (Pi) بسرعة كبيرة.
يتطلب فوسفوريلاز الجليكوجين عامل مساعد هو فوسفات بيريدوكسال (هذا الجزيء هو أيضا عامل مساعد ل transaminases): لديه بروتون فسفوري جزئيا فقط (محاط بفوسفات البيرودوكسال ببيئة مسعورة تبرر وجود البروتونات المرتبطة به) . Phosphoryl (Pi) قادر على إنتاج بروتون للجليكوجين لأن هذا الفسفوريل يستعيده ، البروتون من الفوسفوريل المحمي جزئياً لفوسفات البيروديكسال. إن احتمال أن يفقد الفوسفور البروتون ويخرج بروتوناً بالكامل منخفضاً جداً عند درجة الحموضة الفسيولوجية.
دعونا نرى الآن كيف يعمل الفوسفوغلوكوتاز. هذا الإنزيم له بقايا سينية فسفورية في الموقع الحفزي. ينتج الفسيفوريل إلى فوسفور الجلوكوز 1 (في الموضع السادس): لفترة قصيرة ، يتكون الجلوكوز 1،6-بيسفوسفات ، ثم يتم إعادة تثبيت السيرين عن طريق أخذ الفوسفوريل في الموضع الأول. يمكن أن يعمل mutase phosphoglucus في كلا الاتجاهين ، أي تحويل الجلوكوز 1-فوسفات إلى 6-فوسفات الجلوكوز أو العكس ؛ إذا تم إنتاج 6-فوسفات الجلوكوز ، يمكن إرساله مباشرة إلى تحلل السكر ، إلى العضلات ، أو تحويلها إلى الجلوكوز في الكبد.
إنزيم إنزيم الفوليكول ترانسفيراز (أو بيروفوسفوريلاز الجلوكوز UDP) يحفز تفاعل تحويل الجلوكوز-1-الفوسفات إلى UTP عن طريق مهاجمة الفوسفوريل أ.
الإنزيم الموصوف أعلاه هو بيروفوسفيرلاز: هذا الاسم يرجع إلى حقيقة أن رد الفعل المعاكس للواحد الموصوف للتو هو pyrophosphorylation.
الغلوكوز UDP ، تم الحصول عليه كما هو موضح ، قادر على إطالة سلسلة الجليكوجين ، من وحدة أحادي السكاريد.
من الممكن تطوير التفاعل تجاه تكوين الجلوكوز UDP عن طريق التخلص من منتج هو بيروفوسفات ؛ يحول إنزيم بيروفسفاتاز البيروفوسفات إلى جزيئين من أورثوفوسفاتي (التحلل المائي لانهيدريد) وبذلك يبقي تركيز البيروفوسفات منخفضًا إلى حد كبير بحيث يتم تعزيز عملية تكوين الجلوكوز UDP.
كما ذكرنا ، فإن الجلوكوز UDP ، بفضل عمل سينسيز الجليكوجين ، قادر على إطالة سلسلة الجليكوجين.
ترجع النتائج (بنسبة 1:10) إلى حقيقة أنه عندما تتكون سلسلة الجلايكوجين من 20 إلى 25 وحدة ، يتدخل إنزيم متفرع (يمتلك أيستيدين في موقعه الحفاز) ، قادرًا على نقل سلسلة من 7 -8 وحدات glycosidic المصب من 5-6 وحدة: وبالتالي يتم إنشاء فرع جديد.
بالنسبة للأمور ذات المنشأ العصبي أو إذا كانت الطاقة مطلوبة بسبب المجهود البدني ، يتم إفراز الأدرينالين من الغدد الكظرية.
الخلايا المستهدفة من الأدرينالين (والنورادرينالين) هي خلايا الكبد والعضلات والأنسجة الدهنية (في هذا الأخير هناك تدهور في الدهون الثلاثية ودورة الأحماض الدهنية: في الميتوكوندريا ، وبالتالي ، تنتج الجلوكوز 6 -الفوسفات ، ليتم إرساله إلى التحلل السكري ، في حين أنه في الخلايا الشحمية ، يتحول الجلوكوز 6-الفوسفات إلى الجلوكوز عن طريق إنزيم الفوسفاتيز 6-الفوسفات الجلوكوز ويتم تصديره إلى الأنسجة).
دعونا نرى ، الآن طرق عمل الأدرينالين. يذهب الأدرينالين للربط لمستقبل يقع على غشاء الخلية (من الخلايا العضلية وخلايا الكبد) وهذا يحدد ترجمة الإشارة من الخارج إلى داخل الخلية. يتم تنشيط بروتين كيناز الذي يعمل في وقت واحد على الأنظمة التي تنظم توليف الجليكوجين وتدهوره:
يتواجد سينزاس الجليكوجين في شكلين: شكل مفكك (نشط) وشكل فسفوريليته (غير نشط) ؛ بروتين كيناز فسفوريليز الجليكوجين سينسيز وكتل عملها.
يمكن أن يوجد فوسفوريلاز الجليكوجين في شكلين: شكل نشط يوجد فيه سيرين مفكك و شكل غير نشط يتم فيه فصل السرين. يمكن تنشيط الفوسفوريلاز الجليكوجين عن طريق انزيم فوسفوريلاز جلايكوجين الجليكوجين . ينشط الجلايكوجين فوسفوريلاز كيناز إذا كان فسفوريليد وغير نشط إذا كان dephosphorylated؛ بروتين كيناز له ركيزة له كغاز فوسفوريلاز الجلايكوجين ، أي أنه قادر على الفسفرة (وبالتالي ، تنشيط) الأخير الذي ، بدوره ، ينشط فسفوريلاز الجليكوجين.
بمجرد الانتهاء من إشارة الأدرينالين ، يجب أن تنتهي أيضًا من تأثيرها في الخلية: تتدخل إنزيمات الفوسفاتيز ، بعد ذلك ، على الأنواع البروتينية.